Определение перевода
Перевод относится к процессу создания белков из мРНК шаблон. Последовательность нуклеотидов на РНК транслируется в аминокислотную последовательность белков, и эта реакция осуществляется рибосомами. Рибосомы и тРНК состыковать зрелую транскрипт мРНК и привлечь несколько ферментов в энергоемкий процесс, который использует АТФ, а также ГТФ.
Генетический код
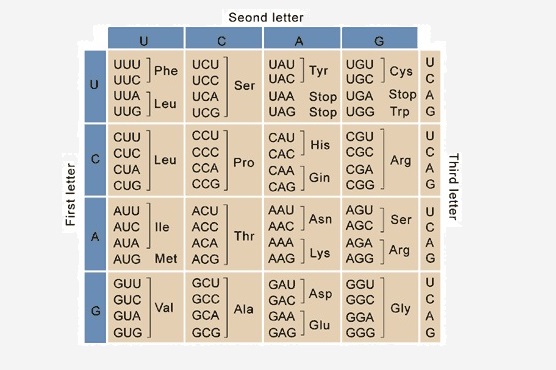
Когда в качестве первичного генетического материала были обнаружены нуклеиновые кислоты, на первый план вышел один важный вопрос. В нуклеиновых кислотах всего 4 основания, тогда как белки состоят из 20 аминокислоты, Следовательно, невозможно иметь прямую взаимно-однозначную корреляцию между последовательностями нуклеотидов и аминокислот. Даже наличие двух нуклеотидов для одной аминокислоты недостаточно. Поэтому было высказано предположение, что аминокислоты кодируются отрезками из трех нуклеотидов, называемых кодонами. Серия экспериментов в 1960-х годах подтвердила эту гипотезу, а также показала, что эти кодоны не перекрываются друг с другом. Кроме того, поскольку отрезки из 3 нуклеотидов могут давать в общей сложности 64 кодона, одна аминокислота может кодироваться несколькими кодонами, свойство, которое называется «вырожденность». Часто разница между вырожденными кодонами – это третье основание, которое называется «положение колебания». Например, аминокислота серин может кодироваться шестью кодонами, четыре из которых: UCA, UCG, UCU или UCC. Точно так же фенилаланин может быть представлен либо UUU, либо UUC на мРНК, а лейцин кодируется в общей сложности шестью кодонами. Этому вырождению способствует тот факт, что третий нуклеотид в каждом кодоне слабо связывается с его соответствующей тРНК, позволяя необычным типам оснований спариваться друг с другом.
Из 64 кодонов, образованных различными комбинациями из 4 нуклеотидов, 3 являются стоп кодоны, что означает конец перевода. Это UAA, UAG и UGA, которые распознаются белками, называемыми факторами высвобождения, а не тРНК. Когда рибосома сталкивается с стоп кодон он отделяется от мРНК благодаря ферментативному действию факторов высвобождения.
мРНК нетранслируемые регионы
Весь участок зрелой мРНК не состоит из кодонов, которые транслируются в аминокислотную последовательность белка. На 5′-конце РНК имеется «крышка», два коротких отрезка нетранслируемых регуляторных областей, примыкающих к кодирующей последовательности (5′-UTR и 3′-UTR), и полиаденилатный хвост, который может определять последовательность белка без прямой трансляции ,

Эти регионы участвуют в экспорте мРНК из ядра, защите от ферментативной деградации и регуляции трансляционной активности. Они содержат сайты связывания белков, которые могут усиливать или уменьшать трансляцию, сайты стыковки рибосом и других частей механизма трансляции, а также ферменты, которые катализируют деградацию мРНК, когда потребность белка в белке клетка встретил
Структура и функции тРНК
Трансферная РНК действует как адаптерная молекула между мРНК и аминокислотами, доставляя соответствующую аминокислоту в рибосому на основе кодонов мРНК. тРНК содержат три основания антикодоновую который может распознавать и связываться с мРНК, а также действовать как сигнал для правильной аминокислоты. Последовательность антикодона является комплементарной кодону мРНК и проходит в антипараллельном направлении, что позволяет двум молекулам базовая пара друг с другом.
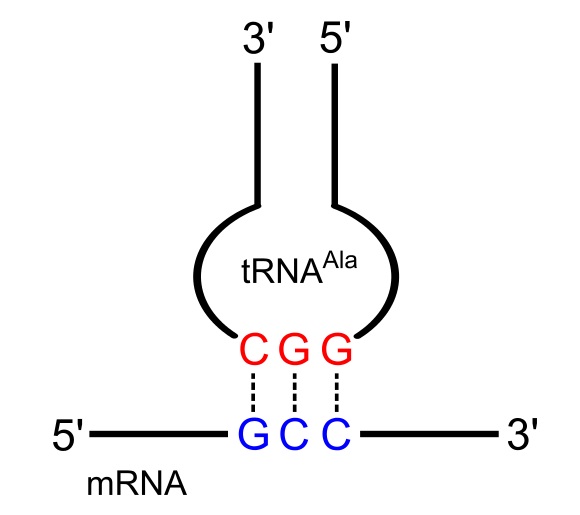
Группа ферментов, называемых аминоацил-тРНК-синтетазой, присоединяет соответствующую аминокислоту к молекулам тРНК на основе их антикодона. Существует одна аминоацил-тРНК-синтетаза для каждой из 20 аминокислот, и фермент может распознавать все антикодоны, которые представляют эту конкретную аминокислоту. Эти ферменты используют энергию АТФ для присоединения аминокислоты к последнему нуклеотиду на 3′-конце тРНК. Теперь считается, что тРНК «заряжена» и может участвовать в реакциях синтеза белка на рибосоме.
Структура и функция рибосомы
Рибосомы представляют собой макромолекулярные мультисубъединичные структуры, содержащие как РНК, так и белок, и являются основными механизмами, которые управляют синтезом белка. Структура рибосомы происходит в первую очередь от его РНК-компонента (рибосомная РНК или рРНК) и спаривание оснований с мРНК и тРНК имеет решающее значение для его функции.
Рибосома содержит две субъединицы, и трансляция инициируется, когда меньшая субъединица связывается с последовательностями выше кодирующей последовательности на мРНК. Прокариотическая трансляция начинается с того, что рРНК непосредственно связывается с мРНК, тогда как эукариотическая трансляция включает другие белки, называемые факторами инициации. Меньшая субъединица вместе с некоторыми другими белками рекрутирует большую субъединицу рибосомы, и начинается трансляция.
Прежде всего, рибосома содержит три важных области – сайт P, сайт A и сайт E -, образованные трехмерной формой рРНК. Сайт P связывается с растущим полипептидом, сайт A закрепляет входящую заряженную тРНК и после пептидная связь образование, тРНК кратко связывается с сайтом Е, прежде чем покинуть рибосому.
Механизм перевода
Перевод осуществляется в три этапа – инициация, удлинение и окончание. Каждый из них связан с различными белками, и на каждом этапе АТФ и ГТФ используются в качестве источников энергии.

Одна мРНК может транслироваться несколькими рибосомами в процессе, называемом транслатомой. Эти комплексы первоначально назывались эргосомами и теперь называются полисомами или полирибосомами.
инициирование
Трансляция начинается с того, что зрелый транскрипт мРНК экспортируется из ядра, а его 5′-шапка распознается меньшей субъединицей рибосомы. Рибосомная субъединица вместе со специальной тРНК сканирует мРНК, чтобы найти начальный сайт для трансляции, который часто является AUG – кодоном для метионина. Последовательности вокруг AUG стартовый кодон также важны и могут определить, насколько сильно транслируется мРНК. Инициирование также включает в себя активность ряда белков-помощников, называемых факторами инициации, функция которых состоит в том, чтобы гарантировать, что различные части механизма трансляции собираются вместе упорядоченным образом.
Рибосома и инициаторная тРНК медленно движутся вдоль мРНК, пока не будет найден стартовый кодон. Структурные особенности этой тРНК гарантируют, что она распознается факторами инициации и распознается факторами элонгации трансляции. Метионин, присоединенный к этой тРНК, также приспособлен для использования исключительно в качестве инициатора – аминогруппа модифицирована для образования N-формилметионина, предотвращая его участие в фазе удлинения трансляции.
Как только большая субъединица рибосомы прибывает в сайт начала трансляции, и тРНК располагается в ее Р-сайте, инициация считается завершенной.
относительное удлинение
Связывание инициатора тРНК с Р-сайтом на рибосоме приводит к ее тесному контакту с амино- или «А» -символом рибосомы, где следующий кодон ожидает трансляции. Новые тРНК, несущие аминокислоты, поступают в рибосому на сайте А. Посредством дополнительного сопряжения оснований и энергии от одного молекула GTP, правильная тРНК связывается с рибосомой. Рибосомная РНК затем катализирует образование пептидной связи между первой и второй аминокислотами, где первый метионин, по-видимому, «перенес» в тРНК на сайте A. Как только пептидная связь образуется, пустая тРНК выходит из рибосомы, и сама рибосома движется вперед ровно на один кодон, так что тРНК на А-сайте перемещается в Р-сайт. Это также подвергает следующий кодон для сайта А, готовый к третьей тРНК. Этот процесс продолжается до тех пор, пока не будет достигнут стоп-кодон.
![]()
прекращение
Когда стоп-кодон присутствует в А-сайте, он распознается рядом белков, называемых факторами высвобождения. Они побуждают рибосому присоединять молекулу воды к растущей полипептидной цепи, а не другую аминокислоту. Это прекращает процесс трансляции и высвобождает полипептид из рибосомы. Две субъединицы рибосомы также отделяются друг от друга, готовые к следующему циклу трансляции.
Перевод на эндоплазматический ретикулум
Трансляция может происходить либо на свободных рибосомах в цитоплазма или на рибосомах, присутствующих на поверхности эндоплазматическая сеть (ЭР). Трансляция начинается в цитоплазме почти для всех белков. Однако белки, которые необходимы в качестве белков внутренней мембраны или те, которые должны секретироваться из клетки, нацелены для дальнейшей трансляции на ER. Эти белки содержат короткий участок гидрофобный остатки называют сигнальным пептидом в начале их последовательности. Как только эти остатки транслируются, сигнальный пептид распознается специфическими белками, называемыми частицами распознавания сигнала, которые могут транспортировать всю рибосому и связанные молекулы к мембране ER. Сигнальный пептид внедряется в мембрану ER, а остальная часть белка высвобождается во внутреннее пространство ER. Сигнальный пептид удаляется из белков, которые должны секретироваться из клетки, тогда как те, которые предназначены для внутренних мембран, сохраняют этот короткий участок, обеспечивая мембранный якорь.
Иногда белки, необходимые внутри органелл, таких как митохондрии и хлоропласты транслируются в цитоплазме. Эти белки селективно транспортируются в эти органеллы с использованием специфических белков в энергоемком процессе.
Антибиотик мишени
Различия между прокариотическим и эукариотическим механизмом перевода делают их идеальными лекарственными мишенями для борьбы с инфекциями, оставляя при этом клетки организма нетронутыми. Эти антибиотики включают хлорамфеникол, тетрациклин, пуромицин и эритромицин. Тем не менее, так как большинство животных также содержат богатый внутренний биом симбиотического бактерии Эти антибиотики также могут вызывать ряд побочных эффектов, включая дефицит витаминов.
- 3 ′ -> 5 ′ ориентация – Направленность одной цепи нуклеиновая кислота которая происходит от нумерации атомов углерода на нуклеотидном сахарном кольце. Один конец нуклеиновой кислоты имеет свободный гидроксильная группа на третьем углероде, а другой конец имеет свободный фосфатная группа прикреплен к пятому карбону.
- Антипараллельная ориентация – Два биополимера, которые идут параллельно друг другу, но в противоположных ориентациях. Две цепи ДНК антипараллельны друг другу в отношении ориентации их сахарофосфатных остовов.
- Энергетическая валюта клетки – Маленькие нуклеотиды, содержащие высокоэнергетические связи, которые используются для хранения и высвобождения энергии внутри клетки. АТФ и GTP являются общими энергетическими валютами в клетке.
- Шероховатой эндоплазматической сети – эндоплазматический ретикулум, усеянный рибосомами, где белки предназначены для внутренних мембран или для секреция переведены.
викторина
1. Какой из этих кодонов кодирует серин?A. UCAB. UAAC. UAGD. Все вышеперечисленное
Ответ на вопрос № 1
верно. Хотя серин может кодироваться несколькими кодонами, четыре из них отличаются друг от друга только третьей позицией или позицией колебания. Следовательно, UCA, UCC, UCG и UCU кодируют серин. UAA и UAG являются стоп-кодонами.
2. Какой из этих ферментов участвует в трансляции?A. Аминоацил тРНК-синтетазыB. 23S рРНКC. ПептидилтрансферазаD. Все вышеперечисленное
Ответ на вопрос № 2
D верно. Аминоацил-тРНК-синтетаза участвует в зарядке тРНК соответствующей аминокислотой. Большая субъединица прокариотической рибосомы содержит 23S рРНК, которая действует как фермент, включая активность переноса пептидила, которая важна для удлинения полипептидной цепи.
3. Сколько аминоацил-тРНК-синтетаз существует в клетке?A. 64 – по одному на каждый кодонB. 61 – по одному на каждый кодон, исключая стоп-кодоныC. 20 – по одной на каждую аминокислотуD. 1 – один фермент катализирует все реакции
Ответ на вопрос № 3
С верно. Отдельная аминоацил-тРНК-синтетаза может распознавать все антикодоны конкретной аминокислоты и заряжать тРНК соответствующим образом.