Определение транскрипции
Транскрипция относится к первому шагу ген выражение, где полимер РНК создается из матрицы ДНК. Эта реакция катализируется ферментами, называемыми РНК-полимеразами, и РНК-полимер антипараллелен и комплементарен матрице ДНК. Участок ДНК, который кодирует РНК-транскрипт, называется единицей транскрипции и может содержать более одного гена.
Эти РНК-транскрипты могут либо использоваться в качестве мессенджеров для управления синтезом белков, либо участвовать в ряде различных клеточных процессов. Эти функциональные или некодирующие РНК могут быть переносимыми РНК (тРНК ), рибосомная РНК (рРНК), или прямая регуляция генов посредством интерференции РНК (RNAi) и формирования гетерохроматина.
Функция транскрипции
Говорят, что жизнь на Земле началась с самореплицирующейся РНК, поскольку это единственный класс молекул, способных как к катализу, так и к переносу генетической информации. С развитием белки взяли на себя катализ, потому что они способны к большему разнообразию последовательностей и структур. Кроме того, связи на сахарофосфатном остове РНК уязвимы даже для незначительных изменений pH и могут подвергаться щелочному гидролизу. Таким образом, ДНК стала предпочтительной молекула для переноса генетической информации, поскольку она более стабильна и устойчива к деградации. Транскрипция поддерживает связь между этими двумя молекулами и позволяет клеткам использовать стабильный нуклеиновая кислота как генетический материал, сохраняя при этом большую часть своего механизма синтеза белка.
Кроме того, отделение ДНК от сайта для синтеза белка также защищает генетический материал от биохимических и биофизических стрессов сложных, многослойных процессов. Небольшие ошибки в РНК-транскрипте могут быть преодолены, поскольку молекула РНК имеет короткий период полураспада, но изменения в ДНК становятся наследственными мутациями. Кроме того, транскрипция добавляет еще один слой для сложной генной регуляции, особенно в вид с большими геномами, которые требуют незначительной корректировки метаболизма.
У эукариот транскрипция также играет важную роль в передаче информации от ДНК к цитоплазма потому что ядерная пора слишком мала, чтобы пропустить рибосомы, белки или хромосомы. В то время как ядерная пора имеет диаметр около 5-10 нм, рибосомы имеют размер от 25 до 30 нм, многие белки имеют ширину более 10 нм, а полностью конденсированные хромосомы могут иметь размер более 2000 нм. Следовательно, основной механизм синтеза белка не может проникнуть в ядро, а отрезки ДНК не могут выйти из ядра.
Механизм транскрипции
Транскрипция создает одноцепочечную молекулу РНК из двухцепочечной ДНК. Поэтому только информация в одной из цепей передается в нуклеотид последовательность РНК. Одна нить ДНК называется кодирующей нитью, а другая – матричной нитью. Механизм транскрипции взаимодействует с шаблоном, чтобы произвести мРНК чья последовательность напоминает кодирующую цепь. Другие названия для шаблонной цепи включают антисмысловую цепь и главную цепь.
![]()
Два разных гена на одной молекуле ДНК могут иметь кодирующие последовательности на разных цепях.
Транскрипционная активность особенно высока в фазах G1 и G2 клеточный цикл когда клетка либо восстанавливается после митоз или готовится к драматическим событиям следующего цикла деление клеток.
Инициация транскрипции
Транскрипция начинается со связывания RNAP в присутствии общих факторов транскрипции с промоторной областью перед сайтом начала транскрипции на ДНК. Прокариотическая РНКП связывается с сигма-фактором, тогда как эукариотическая РНК-полимераза может взаимодействовать с рядом факторов транскрипции, а также с белками-активаторами и репрессорами. Первоначально после связывания RNAP с областью промотора ДНК остается в двухцепочечной форме. Это называется «замкнутый комплекс» между ДНК и RNAP. После этого RNAP вместе с факторами транскрипции разматывает сегмент ДНК и взаимодействует с открытыми нуклеотидами в открытом комплексе, создавая «пузырь транскрипции». RNAP затем отправляется в путешествие по ДНК, сканируя сайт начала транскрипции внутри пузыря. Как только начальный сайт расположен, первые два нуклеотида транскрипта связываются друг с другом.
Побег из Промоутер
После добавления первых нескольких нуклеотидов в предполагаемый РНК-транскрипт РНКП вступает в критическую нестабильную фазу. Он может либо продолжать к продуктивной инициации, либо притягивать ДНК к себе, создавая сжатую открытую ДНК внутри полимеразы. Если RNAP перематывает нижнюю часть ДНК, предполагаемая РНК-транскрипт высвобождается, потому что комплекс ДНК-RNAP возвращается к своей первоначальной открытой конфигурации. Это называется неудачной инициацией.
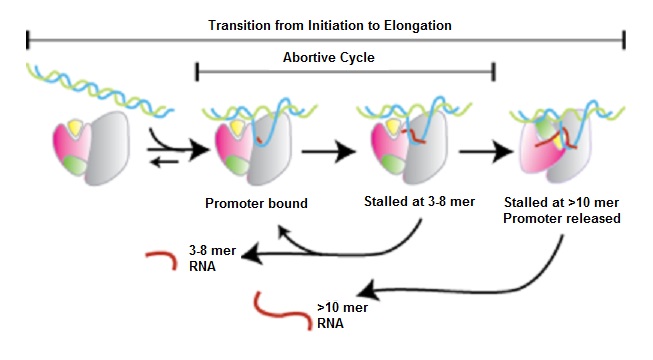
Однако, если верхняя часть ДНК перематывается и выталкивается из фермента, RNAP продвигается вперед. Его взаимодействие с промоторной областью нарушается, и РНК-транскрипт достигает длины 14-15 нуклеотидов. Это называется побегом от промотора и сопровождается изменениями во взаимодействиях белок-белок и белок-ДНК. Некоторые транскрипционные факторы высвобождаются, и транскрипция движется к фазе удлинения.
Удлинение транскрипции
Как только образуется короткий РНК-олигонуклеотид из более чем 15 оснований, РНКП проходит вдоль цепи матричной ДНК. Стенограмма идентична кодирующей цепи, за исключением того, что в нуклеотидном остове вместо сахара содержится рибоза дезоксирибоза и пары оснований аденина с урацилом вместо тимин, RNAP может катализировать образование фосфодиэфирной связи между пятым атомом углерода входящего нуклеотида и третьим атомом углерода последнего нуклеотида в существующем транскрипте.

Поскольку молекула РНК имеет свободный фосфат, присоединенный к пятому углероду на первом нуклеотиде, и свободный гидроксильная группа говорят, что на третьем углероде последнего нуклеотида РНК транскрибируется в направлении от 5 ‘до 3’.
Завершение транскрипции
В отличие от репликации ДНК, где ДНК-полимераза продолжает добавлять нуклеотиды, пока не достигнет конца молекулы, транскрипция должна быть прекращена в определенном месте для эффективной регуляции и экспрессии генов. Прекращение прокариотической транскрипции может происходить через образование двухцепочечной области внутри РНК или под действием белка, называемого Rho.
Первый метод включает транскрипцию богатой G: C области, за которой следует цепочка урацилов, которые образуют слабые водородные связи с матричной ДНК. Обогащенная G: C область может зацикливаться на себе, образуя шпилеподобную структуру, блокирующую RNAP и механизм транскрипции. Это в сочетании с более слабыми связями между урацилом и матричной ДНК может отодвинуть РНК от механизма транскрипции и привести к терминации. Этот процесс также включает белок под названием NusA.

Rho-зависимая терминация транскрипции включает связывание белка Rho с последовательностью на транскрибируемой РНК. Эта последовательность, которая находится ниже по течению от перевод стоп кодоны Позволяет Rho связываться с РНК и перемещаться по транскрипту АТФ-зависимым образом. Когда он сталкивается с остановленным RNAP, он связывается с ферментом и заставляет транскрипт и связанный с ним механизм диссоциироваться от ДНК.
Прекращение эукариотической транскрипции гораздо менее изучено, и большая часть работы была сосредоточена на механизмах RNAP II. Терминация транскрипции у эукариот также связана с посттранскрипционными модификациями и процессингом перед экспортом зрелой РНК в цитоплазму.
Типы РНК-транскриптов
Традиционно были известны три типа транскриптов РНК – мессенджер РНК (мРНК), тРНК и рРНК – и все три тесно связаны с синтезом белка. Хотя мРНК определяет аминокислотную последовательность, тРНК и рРНК имеют решающее значение для механизма трансляции кода мРНК.
Полимеризация мРНК из ДНК-содержащих белков, кодирующих гены, катализируется РНК-полимераза II. Иногда белки, которые используются вместе, кодируются как единое целое в одной длинной молекуле мРНК, и это особенно распространено среди прокариот. Последовательности ДНК перед кодирующей последовательностью содержат сайты стыковки для механизма транскрипции, а также регуляторные факторы, которые модулируют время и количество транскрипционной активности. Затем мРНК модифицируют и обрабатывают для получения окончательного транскрипта, используемого для трансляции.
рРНК составляет почти пятьдесят процентов РНК клетки и транскрибируется РНК-полимеразой I в специализированных областях ядра, называемых ядрышками. Ядрышки представляют собой плотные сферические структуры вокруг локусов, кодирующих рРНК. Прокариотическая рРНК имеет три типа, а эукариотические рибосомы состоят из четырех типов рРНК, самая большая из которых содержит более 5000 нуклеотидов. Эти молекулы РНК определяют трехмерную структуру рибосом.
РНК-полимераза III катализирует транскрипцию предшественников тРНК в ядре. Промоторные последовательности, контролирующие экспрессию генов тРНК, могут быть внутригенными, расположенными внутри кодирующей последовательности гена. Предшественники тРНК подвергаются обширным модификациям, включая сплайсинг. Прокариотические тРНК сохраняют свою каталитическую активность и могут самостоятельно сплайсироваться, тогда как эукариотическая посттранскрипционная модификация осуществляется специальными эндонуклеазными ферментами. Эти эндонуклеазы распознают специфические структурные мотивы в тРНК, которые нацелены на последовательность для сплайсинга.
В дополнение к этим трем типам РНК клетка содержит ряд более мелких РНК, участвующих в различных клеточных активностях. К ним относятся генная регуляция (опосредованная микроРНК и последовательностями в 5′-нетранслируемых областях транскриптов мРНК), посттранскрипционная модификация (малая ядерная РНК, малая ядрышковая РНК), защита генома (Piwi-взаимодействующая РНК и CRISPR ) и поддержание геномной структуры (теломеры и РНК-транскрипты, которые молчат Х-хромосомы).
Различия между прокариотической и эукариотической транскрипцией
Очевидная разница между прокариотической и эукариотической транскрипцией заключается в наличии ядерная мембрана у эукариот. Эукариотические РНК-транскрипты необходимо экспортировать из ядра, тогда как прокариоты осуществляют сопряженную транскрипцию и трансляцию в цитоплазме. Это возможно, потому что прокариотический транскрипт не подвергается обширной модификации, а прокариоты не нуждаются в транскрипционных факторах для инициации. Следовательно, механизм транскрипции проще и может одновременно вместить ферменты трансляции.
Прокариоты также имеют только одну РНК-полимеразу, чтобы катализировать все реакции транскрипции клетки, и один РНК-транскрипт может направлять синтез нескольких белков. Эти мРНК называются полицистронными мРНК. Часто все гены, участвующие в одном биохимическом пути, транскрибируются и транслируются вместе, что позволяет регулировать весь путь как единое целое. У эукариот поликистронная мРНК может быть обнаружена в хлоропластах.
- Моноцистронная мРНК – транскрипт мРНК, кодирующий отдельный белок.
- Транспозоны – Небольшие сегменты ДНК, которые могут перемещаться по геному, вставляя себя в локусы, удаленные от их исходного сайта, часто с участием РНК-интермедиата.
- hnRNA – Гетерогенные ядерные РНК считаются исходными продуктами транскрипции и состоят в основном из предшественников мРНК.
- Поли-А-полимераза – Фермент, который добавляет участок адениннуклеотидов к концу первичного транскрипта.
викторина
1. Какие из этих свойств делают ДНК более стабильным генетическим материалом?A. Водородные связи между основаниями сильнееB. ДНК длиннее РНКC. Наличие тиминовых основанийD. Стойкость к деградации в результате щелочного гидролиза
Ответ на вопрос № 1
D верно. Сахарная цепь ДНК содержит дезоксирибозу, что делает ее более устойчивой к деградации в результате щелочного гидролиза. Водородные связи не особенно прочны в ДНК по сравнению с РНК, несмотря на изменения в базовых составах.
2. Каков размер ядерной поры у эукариот?A. Менее 10 нмB. Более 10 нмC. Более 2000 нмD. 25-30 нм
Ответ на вопрос № 2
верно. Ядерные поры имеют размер 5-10 нм. Многие белки имеют размер более 10 нм, размер рибосом составляет 25-30 нм, а полностью конденсированных хромосом – более 2000 нм. Ядерные поры действуют как эффективные сита, которые контролируют движение макромолекул в ядро и из него.
3. Что из этого НЕ является особенностью экспрессии генов прокариот?A. Сопряженная транскрипция и переводB. Обширная посттранскрипционная модификация РНК-транскриптаC. Сигма-фактор для инициации транскрипцииD. Ни один из вышеперечисленных
Ответ на вопрос № 3
В верно. Отсутствие ядерной оболочки позволяет связывать прокариотическую транскрипцию и трансляцию, а транскрипция инициируется сигма-факторами. Однако, в отличие от экспрессии гена эукариот, РНК-транскрипт у прокариот имеет меньше интронов и не подвергается значительной модификации.